Человек с хвостом, Парень шокировал хвостом на спине — ТСН, новости 1+1 — Курьезы
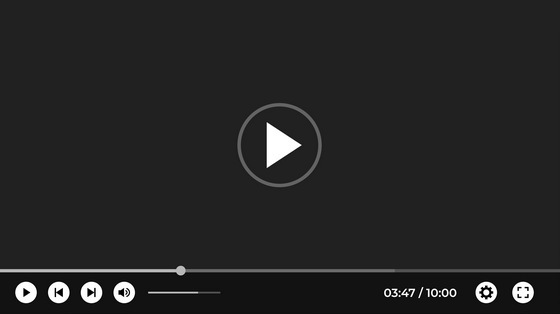
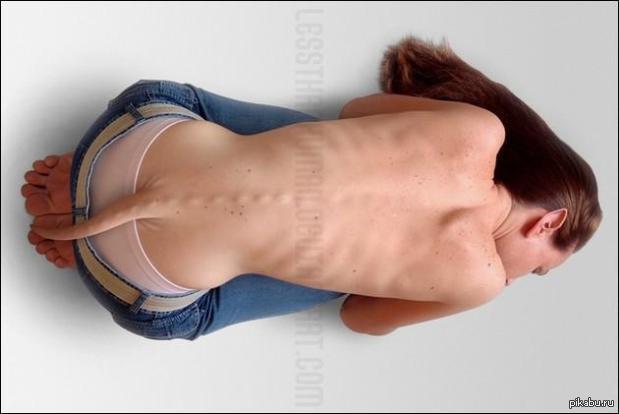
По видимому, выигрыш от этого был достаточно крупным, чтобы перевесить вред, поскольку та же самая мутация может вызывать дефекты спинного мозга, наблюдаемые у одного из тысячи человеческих младенцев. Выявление сдавления структур конского хвоста, отек корешков конского хвоста является показанием к экстренной операции. Результат д.
И это напомнило мне о моем давнишнем интересе. Так и началась эта работа». Сравнив геномы гоминоидных и негоминоидных приматов, и убедившись, что ни один из 31 гена-кандидата у гоминоидов не содержит в своих экзонах мутаций, которые должны были бы как-то заметно нарушить функции белков, кодируемых этими генами, ученые взялись за более внимательное изучение некодирующих участков — в частности, интронов для этих же генов. Общим для всех гоминоидов отличием оказалась вставка мобильного элемента AluY в интрон, разделяющий 6 и 7 экзоны гена TBXT T-box transcription factor T.
Этот белок связывается с регуляторными участками около множества разных генов, и влияет на интенсивность транскрипции этих генов. Собственно, свое название Brachyury от лат.
Мутация давала короткохвостых мышей только при гетерозиготном генотипе, в то время как гомозиготный генотип был летален для эмбрионов. Мутации в этом же гене обуславливают и короткохвостость некоторых пород кошек. Гены всех эукариот живых существ, клетки которых содержат ядро , а значит и всех животных, состоят из экзонов и интронов.
На молекуле РНК далее синтезируется белок. Но между этими двумя этапами считывания генетической информации должно произойти еще кое-что: из первичной РНК должны быть удалены «бессмысленные» участки не кодирующие белок — интроны, а остальные, кодирующие — то есть экзоны — соединяются снова в цельную цепочку, образуя зрелую мРНК, кодирующую полноценный белок.
Этот процесс называется сплайсингом. Надо иметь в виду, что зачастую сплайсинг может происходить несколькими разными способами, что приводит к образованию нескольких разных вариантов зрелой мРНК, а значит и нескольких вариантов белка при последующей трансляции. Очевидно, что внутриклеточная молекулярная машина, сплайсосома , которая выполняет эту операцию, должна каким-то образом опознавать, где именно надо произвести вырезание эти места именуются «сайты сплайсинга».
Действительно, внутри генов, как в экзонах, так и в интронах есть особые нуклеотидные последовательности, которые выполняют роль опознавательных и направляющих знаков для этой машины. Кроме того, важно, какую пространственную конфигурацию принимает молекула первичной РНК. Сайты для посадки сплайсосомы в некоторой конфигурации могут оказаться как бы спрятаны от сплайсосомы, и тогда сплайсинг будет проходить альтернативным путем, не задействуя скрытые сайты сплайсинга.
Вернемся к обнаруженной мутации. На что же может влиять мобильный элемент, оказавшийся в середине интрона?
Предположили, что он влияет на сплайсинг. Как же именно? Причем направлен он в противоположную сторону по отношению к новому элементу AluY генома гоминоидов. Это взаиморасположение теоретически может приводить к комплементарному спариванию двух Alu-элементов в процессе транскрипции с образованием петли, содержащей шестой экзон, отделенной от всей остальной части цепочки РНК «стволом» из пары мобильных элементов.
РНК, которая получается при таком варианте сплайсинга, оказывается короче полного варианта. Теряющийся фрагмент не нарушает рамку считывания , но приводит к образованию укороченного на 59 аминокислот белка с выпадением одного трансактиваторного TA1 и одного репрессорного домена TR1 — это показано на рис. Таким образом, утрата этого экзона, очевидно, должна сказываться на функциональности альтернативной изоформы белка. Теоретический сценарий изменения получаемых продуктов гена при вставке мобильного элемента AluY в интрон из-за альтернативного сплайсинга.
В итоге получаем два разных фенотипа. Теперь дело осталось за экспериментальной проверкой. Первая серия экспериментов была выполнена на культивируемых эмбриональных стволовых клетках человека. Задача — убедиться, что альтернативный сплайсинг действительно присутствует, и — если да, — что причиной действительно является взаимодействие между двумя Alu-элементами. С их помощью в ДНК клеток можно было вырезать один или второй Alu-элемент рис. Получив клетки с такими делециями в геноме каждый вариант в двух независимых повторах , ученые исследовали набор молекул зрелых мРНК гена TBXT , которые в этих клетках образуются.
Результаты можно видеть на рис. Каждая полосочка на электрофорезе — один фрагмент определенной длины. Подписи указывают, какой фрагмент происходит из какого варианта. Очевидно, это элемент задействован не в одном варианте альтернативного сплайсинга. Таким образом, гипотетический молекулярный механизм влияния новой вставки мобильного элемента в интрон гена на альтернативный сплайсинг в полной мере подтвердился. Теперь осталось проверить вторую часть гипотезы — что именно появление такого варианта альтернативного сплайсинга и есть причина укорочения хвоста.
Тут уже не обойтись без экспериментов на животных. Любимый модельный объект в таких случаях — мышь. Но вот беда, у мышей вовсе нет никаких Alu-элементов: это чисто приматное семейство мобильных элементов. Что же делать? При их скрещивании во втором поколении теоретически можно получить мышей, имеющих гомозиготный нормальный генотип, а также гетерозиготный и гомозиготный генотип с делецией экзона в соотношении , согласно законам расщепления Менделя.
Однако на практике у мышей наличие мутации в гомозиготе приводило к аномалии развития, так что эмбрион либо погибал на самых ранних этапах развития, либо развивался, но детеныш умирал при рождении из-за патологии в виде расщепления позвоночника. Так что живые мышата получались только в том случае, если могла нарабатываться как полноразмерная, так и укороченная версия белка либо только полноразмерная , — и ведь именно так обстоит дело в случае, когда укороченная изоформа получается путем альтернативного сплайсинга, как у гоминоидов!
Проверка влияния альтернативного сплайсинга на формирование фенотипа с использованием генетически отредактированных мышей. B — схема строения зрелых РНК у мышей с нормальным или гетерозиготным генотипом, а также у человека.
Как и следовало ожидать, мышата второго поколения с двумя нормальными хромосомами имели самый обычный хвост без каких-либо особенностей 35 из А вот мышата с гетерозиготным генотипом сильно разнились.
В целом, все части тела, не считая хвоста, у них имели вполне нормальный вид. Что касается хвоста — то тут спектр фенотипов варьировал от полного отсутствия хвоста до хвоста обычной длины и нормального фенотипа. Весь спектр можно видеть на рис. Аномалии фенотипа присутствовали у 21 из 63 мышат-гетерозигот, у четырех из них хвоста не было совсем no-tail , у 9 хвост был заметно укорочен short-tail — в их хвосте было намного меньше позвонков, чем бывает обычно, а еще у восьми присутствовали аномалии в виде «изломов» хвоста kinked-tail.
Таким образом, присутствие мутации проявляло себя не у всех особей — в таких случаях говорят о «неполной пенетрантности» мутации. Подводя итоги своего исследования, ученые рисуют нам следующий вероятный сценарий того, как предки гоминоидов теряли хвост. Следствием этого стало появление у особей неустойчиво проявляющихся аномалий хвоста — у некоторых это была полная потеря хвоста, у других он мог быть укороченным.
Вероятно, такие изменения дали какое-то преимущество особям, и мутация была поддержана отбором. Со временем нашлись и другие мутации, которые усилили силу влияния мутации на онтогенез и стабилизировали бесхвостый фенотип.
Таких мутаций могло быть намного больше, чем одна, и в рамках этого исследования мы не можем сказать, какие именно гены эти мутации затронули. Увы, такое исследование, раскрывая в значительной мере молекулярный механизм потери хвоста, все еще ничего не может нам сказать о том, почему этот фенотипический признак оказался полезным для наших далеких предков 25 миллионов лет назад. Данные по ископаемым говорят о том, что первые бесхвостые приматы все еще использовали передвижение на четырех лапах и были хорошо адаптированы к жизни на деревьях, так что привязать появление признака к перемещению на задних конечностях не получается.
Будем рады предложению ваших версий в комментариях. Вероятный сценарий хода эволюционных событий, в результате которого у гоминоидов исчез хвост. Интересно, что аномалия в виде расщепления позвоночника довольно-таки нередка у людей частота примерно 1 случай на — новорожденных. Так что, рассуждают авторы, фиксация такой мутации у предкового вида вероятнее всего говорит о наличии какого-то действительно серьезного преимущества, связанного с утратой хвоста для тех животных в их местообитании.
Ну и с другой стороны, это изменение, так или иначе, стало важной преадаптацией, облегчившей переход уже непосредственно предков человека австралопитеков к ходьбе и бегу на двух задних теперь нижних конечностях.
Dasen, Matthew T. Maurano, Sang Y. Kim, Jef D. Boeke, Itai Yanai. DOI: Закрыть Новости науки. Большой адронный коллайдер. Результаты LHC. Загадки LHC. Двухфотонный всплеск ГэВ. LHC в работе.
Общее расписание. Ранние этапы. Результаты работы LHC в году. Сеанс LHC Run 1. Сеанс LHC Run 2.
Сеанс LHC Run 3. Устройство и задачи LHC. Устройство LHC. Задачи LHC. Свойства адронов. Поиск бозона Хиггса. Физика элементарных частиц.
Величины и единицы. Как изучают частицы. Эксперименты на коллайдерах. Стандартная модель. Хиггсовский механизм. В популярных журналах. Как подписаться. Выставка «Всё в мире относительно». Книжный клуб. Опубликовано полностью. Происхождение жизни. Глава 1. Глава 2. Глава 3. Масштабы: времена. В помощь читателю. От секунды до года. Астрономические времена. Фолдинг белков. Возбужденные атомы. Возбужденные атомы: кто такие и где встречаются. Ядерные распады.
Элементарные частицы.
Повышенная жизнеспособность на околосветовых скоростях. Движение континентов. Покорение воздуха. Аппараты легче воздуха. Первый аппарат легче воздуха, способный поднять человека. Типичный дирижабль конца XIX века рыбообразной формы.
Цельнометаллический бескаркасный дирижабль с изменением объема в полете и с подогревом газа. Серийный дирижабль Германии времен Первой мировой войны.
Гибель «Гинденбурга» 6 мая года. Аппараты тяжелее воздуха. Первый вертолет И. Первый летающий аппарат классической схемы. Единственный российский серийный десантный экраноплан. Теоретические основы полета аппаратов тяжелее воздуха. Первый реализованный проект, однако достоверных сведений о полете нет. Лучший истребитель битвы за Британию в г. Цельнодеревянный скоростной бомбардировщик «Мечта термита».
Истребитель — символ японской авиации. Фронтовой истребитель, один из первых серийных реактивных самолетов.
Многоцелевой истребитель, противник МиГ в корейской войне. Стратегический бомбардировщик с велосипедным шасси. Вечный двигатель. Электромагнитное излучение. Возможности человека. Журнал общей биологии. In English. Наука, образование и право. Интеллектуальная собственность.

Поиск Закрыть. Новости науки. Как предки человека остались без хвоста Показать комментарии Свернуть комментарии Сколько же ещё подобных мутаций в "некодирующих" областях остались за кадром до настоящего времени из-за упоротости и пр. Вопрос риторический, если чё Это так при сравнении хвостатых vs бесхвостых?.. Да уж точно, полно 2. Читайте внимательнее. Так что нет, экзон 6 очень консервативным остается и у гоминоидов.
Мысля возникла: можно попробовать засайленсить с помощью иРНК, например у гомозигот. Результат д. И ещё: если удвоить ген или усилить его транскрипцию , то что будет? Хвост позвоночник? Можно точно утверждать, что предковой к гоминоидам обезьяне хвост был совершенно не нужен, иначе бы эта мутантная обезьяна бы не выжила, у нее не было бы потомков. Значит бесхвостые обезьяны ничего не потеряли утратив хвост.
Хвост обезьян как минимум очень энергозатратная вещь, его владение требует больших затрат энергии, так что вполне возможно что бесхвостые обезьяны получили преимущество тем, что смогли перенаправить высвободившуюся энергию, что ранее шла на хвост, на конечности брахиация и мозг. А зачем хвост, например, львам? Почему бы им тоже не "перенаправить энергию"? Львы не обезьяны, их хвост не потребляет энергию, его потеря не изменит баланса энергии, но уменьшит защиту от паразитов и управляемость при беге.
Он состоит из 10 пар спинномозговых нервов и по виду действительно напоминает конский хвост. Синдром конского хвоста -это редкое, но очень серьезное заболевание, причиной которого является выраженное сдавливание компрессия корешков, приводящее к их отеку, нарушению функций и при отсутствии лечения к необратимому повреждению.
При синдроме конского хвоста необходимо проведение экстренной нейрохирургической операции, иначе пациент может остаться на всю жизнь инвалидом. В большинстве случаев синдром конского хвоста характеризуется острым началом и все вышеописанные симптомы проявляются в течение 24 часов.
У небольшой части пациентов синдром конского хвоста развивается в течение нескольких месяцев например, при сдавлении нервных корешков растущей опухолью позвоночника или при быстропрогрессирующем стенозе позвоночного канала. Для подтверждения диагноза синдрома конского хвоста требуется проведение незамедлительной магнитно-резонансной томографии МРТ.
Выявление сдавления структур конского хвоста, отек корешков конского хвоста является показанием к экстренной операции. Только экстренная хирургическая операция декомпрессия нервных корешков позволяет сохранить функцию нервных волокон конского хвоста и привести к полному выздоровлению.
Если после началом заболевания и хирургическим вмешательством прошло более 24 часов возможно необратимое сохранение неврологического дефицита в виде паралича конечностей и тазовых расстройств.
Частые вопросы. Заказать звонок. Заочные консультации. Москва, проезд Березовой рощи, д. Запись онлайн. Главная Что мы лечим Боль в позвоночнике Синдром конского хвоста у человека Синдром конского хвоста у человека.